


Cây trồng bị ảnh hưởng nghiêm trọng bởi đất nhiễm mặn ở nồng độ muối cao. Để tạo ra giống cây trồng chống chịu mặn thì việc tìm hiểu các tính trạng chủ chốt tạo ra tính chống chịu mặn là hết sức quan trọng. Các thông tin được cung cấp tổng hợp từ các nghiên cứu được cập nhật gần đây về cơ chế chống chịu mặn ở thực vật. Các nghiên cứu gần đây chỉ ra rằng cơ chế cảm ứng stress và các thành phần truyền tín hiệu stress có thể giữ vai trò quan trọng trong việc điều hòa các đáp ứng của thực vật đối với stress mặn (nồng độ muối cao trong đất).
Các cơ chế về sự vận chuyển Na+ và các cơ chế khử độc Na+ cũng được thảo luận thêm, các kết quả nghiên cứu về sự biến đổi chromatin ở thực vật dưới điều kiện cây bị stress mặn cũng được thảo luận. Các cố gắng gần đây của các nhà khoa học trong việc chọn tạo các giống cây trồng kháng mặn cũng được đề cập, đặc biệt là ứng dụng chỉ thị phân tử (MAS) và kỹ thuật chồng gene kháng.Các câu hỏi vẫn còn để ngỏ và cần được trả lời để làm phong phú thêm kiến thức về cơ chế chống chịu mặn ở cây trồng.
Đất bị nhiễm mặn và các tác động lên cây trồng
Đất bị nhiễm mặn là một trong những vẫn đề hiện tại thách thức nghiêm trọng nền nông nghiệp các quốc gia trên thế giới. Sự tích tụ muối trong đất canh tác do quá trình tưới tiêu, dẫn nước từ các nguồn chứa muối gốc NaCl và từ sự xâm nhập của nước biển. Gia tăng nồng độ muối trong đất làm giảm khả năng hấp thu nước ở thực vật. Một khi rễ hấp thu một lượng lớn các ion Na+ và Cl- thì các ion này ảnh hưởng xấu đến sự tăng trưởng của cây, các ion Na+ và Cl- làm giảm các quá trình biến dưỡng và hiệu quả của quá trình quang hợp thấp.
Do đó, stress mặn ở thực vật có thể phân ra các giai đoạn như stress cân bằng thẩm thấu ở giai đoạn sớm, stress do tích tụ ion Na+ và Cl-. Hơn thế nữa, thực vật giảm thiểu tác hại của ion Na+ thông qua việc loại bỏ Na+ qua mô lá và sự phân ngăn nhằm quy tụ Na+ trong không bào. Các cây trồng không có khả năng chống chịu mặn sẽ làm giảm năng suất, từ đó dẫn đến thu hẹp diện tích đất canh tác. Sự mất dần diện tích đất có thể canh tác được xem như một thách thức nặng nề đối với ngành nông nghiệp do phải đáp ứng đủ lương thực thực phẩm cho nhân loại đang gia tăng số lượng phi mã (dự đoán dân số thế giới đạt khoảng 9.6 tỉ người vào năm 2050).
Với tầm quan trọng đó, việc tạo ra các giống cây trồng có khả năng chống chịu mặn là chiến lược vô cùng quan trọng để đối phó với thực trạng đất bị nhiễm mặn. Bài tổng hợp này sẽ phân tích các cơ chế chính trong quá trình chống chịu mặn ở thực vật và đưa ra các thông tin cập nhật gần đây về cơ chế cảm ứng stress mặn và sự truyền tín hiệu, cơ chế điều hòa biểu hiện gene ở thực vật. Hơn nữa, việc hiểu rõ mạng lưới vận chuyển ion Na+ trong thực vật và các phương pháp phân tích Na+ cũng giúp ích trong quá trình cải tiến giống cây trồng chống chịu mặn.
Cơ chế cảm ứng stress mặn
Để có cơ chế đáp ứng hiệu quả đối với stress mặn, thực vật tự phát triển khả năng cảm ứng các thành phần siêu thẩm thấu và ion Na+ của quá trình stress mặn. Hai phương thức cảm ứng trên đã chứng minh rằng cơ chế đáp ứng đối với muối NaCl mang nhiều sự khác biệt so với cơ chế đáp ứng với stress thẩm thấu thuần túy. Nồng độ muối cao trong dung dịch đất sẽ tạo ra stress thẩm thấu ở vùng rễ. Cho đến hiện tại thì các cơ chế phân tử về hoạt động của các sensor thẩm thấu ở thực vật và sensor Na+ vẫn cần được được tìm hiểu thêm.
Protein HK1 (Histidine kinase receptor) cho thấy có thể giúp thay thế chức năng sensor Sln1 ở nấm men, các cây Arabiodopsis được biến đổi gene có biểu hiện vượt mức hay làm mất đi chức năng của HK1 mang các kiểu hình liên quan đến tính trạng hạn hán hoặc bị stress thẩm thấu.
Thực vật biểu hiện rất nhiều các đáp ứng sinh lý học khi gặp stress thẩm thấu, tuy nhiên các nghiên cứu gần đây cho thấy rằng trong số các đáp ứng sinh lý học đó thì một số biểu hiện sẽ bị thay đổi ở các cây mang đột biến gene hk1 trong khi các biểu hiện khác không bị ảnh hưởng.
Điều này chứng tỏ rằng vẫn còn có các protein khác đảm nhận vai trò nhận biết tín hiệu stress thẩm thấu trên các cây Arabidopsis bị đột biến gene hk1. Các sensor về thẩm thấu ở thực vật dường như có liên hệ chặt chẽ về hoạt động đối với các kênh Ca++, điều này thể hiện ở việc thực vật biểu hiện sự gia tăng nồng độ ion Ca++ trong tế bào chất chỉ trong vòng vài giây sau khi bị xử lý với NaCl hoặc mannitol.
Các đáp ứng Ca++ có nguồn gốc hoạt động ở rễ và xuất hiện ở rất nhiều dạng tế bào.Kết quả này dẫn đến suy đoán rằng stress thẩm thấu có thể được cảm ứng bởi hoạt động của các kênh Ca++ giai đoạn đầu của chuổi đáp ứng stress.Các kết quả khác cũng chứng minh phương thức hoạt động cảm ứng mang tính cơ học-thẩm thấu.Các đột biến làm ảnh hưởng lên sự phát triển của lớp cutin cũng làm ảnh hưởng đến các quá trình đáp ứng thẩm thấu bao gồm cả hoạt động tổng hợp hormone ABA ở giai đoạn sau của chuổi phản ứng.Lớp cutin tạo ra sự nâng đỡ và bảo vệ vách tế bào cũng như màng sinh chất, bên cạnh đó lớp cutin cũng làm thay đổi khả năng khuếch tán của nước vào bên trong tế bào.
Do đó, thay đổi đặt tính của lớp cutin tế bào cũng làm thay đổi đặc tính cơ học của quá trình stress liên quan đến nước ở tế bào thực vật. Các phân tử hoạt động như thông tin tín hiệu thứ cấp cũng có liên hệ với con đường truyền tín hiệu Ca++, ví dụ như các gốc oxygen hoạt hóa ROS (reactive oxygen species) và các annexin ở Arabidopsis cũng được ghi nhận hỗ trợ các đáp ứng Ca++ liên quan đến cảm ứng NaCl và ROS. Downstream của dòng tín hiệu Ca++ là các kênh kinase sẽ được kích hoạt, các kinase này bao gồm cả CDPK (calcium dependent protein kinase) và CBL (calcineurin B-like protein) liên quan đến các kênh tương tác CIPK (CBL-interacting protein kinase).
Các hoạt động của các kinase này có thể giúp chuyển các tín hiệu thẩm thấu đến các hoạt động tiếp theo của các protein liên quan và sự biểu hiện gene thông qua quá trình phiên mã. Các yếu tố phiên mã (transcription factor = TF) có thể được kích hoạt trực tiếp bởi kênh Ca++/calmodulin và bao gồm cả các yếu tố kích hoạt phiên mã gắn kết với calmodulin CAMTA (calmodulin-biding transcription activator), GTL (GT element-biding like protein) và MYB.
Mặc dù sự gia tăng nhanh chóng nồng độ Ca++ được xem như là đáp ứng chỉ thị đối với stress thẩm thấu nhưng cũng có thể tồn tại các cơ chế cảm ứng thẩm thấu độc lập với con đường tín hiệu Ca++. Phân tích di truyền về các sensor cảm ứng thẩm thấu cũng như cảm ứng Na+ là tiền đề để hiểu rõ các cơ chế cảm ứng này ở giai đoạn sớm khi thực vật gặp các điều kiện stress.
Sự điều hòa biểu hiện gene ở rễ thực vật khi đáp ứng với stress mặn
Các yếu tố phiên mã (transcription factor) có mối liên hệ chặt chẽ với các cơ chế cảm ứng mặn đóng góp vào các hoạt động mang tính đề kháng đối với stress mặn. Các họ gene quy định các yếu tố phiên mã chính trong các đáp ứng với stress mặn như bZIP(basic leucine zipper), WRKY, AP2/ERF (APETALA2/ETHYLENE RESPONSE FACTOR), MYB, bHLH và NAC.
Các yếu tố phiên mã trên chịu trách nhiệm điều hòa sự biểu hiện của các gene ảnh hưởng đến các mức độ chống chịu stress mặn ở thực vật. Để chống lại việc giảm thế nước được gây ra bởi các thành phần mang tính thẩm thấu khi bị stress mặn, các gene liên quan đến các đáp ứng này được kích hoạt biểu hiện ở mức cao như các gene liên quan đến việc hấp thu các ion vô cơ, sự tổng hợp các osmolyte (các phân tử điều hòa tính thẩm thấu của tế bào). Ở một mức độ nào đó, sự điều hòa biểu hiện gene mức phiên mã (transcriptional regulation) liên quan đến các đáp ứng stress mặn ở thực vật được thúc đẩy bởi các thay đổi liên quan đến quá trình sinh tổng hợp các hormone thực vật (phytohormone)
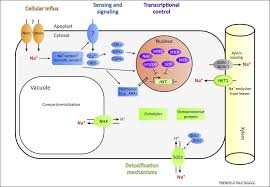
Hình 1: Các cơ chế vận chuyển Na+ và các thành phần quan trọng trong quá trình đáp ứng với stress mặn ở tế bào rễ thực vật
Sau quá trình cảm ứng stress, có một chu kỳ hạn chế các hoạt động tăng trưởng được khởi sự và sau đó là giai đoạn phục hồi. Cả hai giai đoạn này đều liên hệ mật thiết với sự thay đổi lượng hormone ABA (abscisic acid), jasmonate (JA), GA và BR (brassinosteroid). Khai thác nguồn dữ liệu từ AtGenExpress đã khám phá ra hệ thống tín hiệu thứ cấp kiểm soát sự tăng trưởng của thực vật sau quá trình stress mặn.
Trong quá trình đáp ứng với stress độ mặn cao, hầu hết các thay đổi liên quan đến quá trình phiên mã biểu hiện gene xuất hiện chỉ sau khoảng 3h khi cây bị stress mặn. Sự biểu hiện của 5590 gene đã được ghi nhận liên quan đến stress mặn ở bộ phận rễ của cây Arabidopsis thaliana. Sử dụng phương pháp phân biệt các hoạt động riêng biệt của các loại tế bào FACS (fluorescence-activated cell sorting) cho thấy các tế bào vùng lõi của rễ hoạt động phiên mã mạnh nhất.
Các phân tích ở mức phân tử cho thấy vùng nội bì của rễ là lớp tế bào mang chức năng quan trọng đối với sự phát triển của hệ thống rễ phụ dưới điều kiện stress mặn. Hormone ABA ngăn cản sự kéo dài của hệ thống rễ phụ trong môi trường có nồng độ muối cao.
Các nghiên cứu gần đây cũng cho thấy rằng thực vật tránh né môi trường có độ muối cao bằng cách thay đổi hướng tăng trưởng của rễ. Hiện tượng này được giải thích như sự hướng động liên quan đến hiện tượng mặn = halotropism (halo=mặn, tropism=hoạt động liên quan đến yếu tố nào đó). Hiện tượng này không phải do stress thẩm thấu gây ra mà là do các hoạt động đáp ứng với hornmone auxin được gây ra bởi stress mặn. Các cơ chế phân tử về hiện tượng halotropism cần được nghiên cứu thêm để hiểu rõ hơn các cơ chế chống chịu mặn ở thực vật mặc dù có nhiều khám phá quan trọng gần đây về cơ chế chống chịu mặn.
Ngoài các hormone đã được đề cập bên trên, hormone ethylene cũng cho thấy vai trò quan trọng trong sự chống chịu mặn ở cây Arabidopsis thông qua sự thay đổi tỉ lệ Na+/K+ ở chồi. Knock-out (bất hoạt) gene ETO1 (ETHYLENE OVERPRODUCER1) gây ra các thay đổi hàm lượng ethylene và qua đó kích thích quá trình sản xuất ROS ở vùng tủy của rễ bởi yếu tố RBOHF (respiratory burst oxidase homolog F). Sự gia tăng ROS ở vùng tủy dẫn đến giảm dòng ion Na+ vào rễ, giảm thu nhận Na+ ở mạch gỗ (xylem) và duy trì ion K+ ở rễ giúp tăng cường khả năng chống chịu mặn.
Mạng lưới vận chuyển Na+ và K+
Rất nhiều kênh vận chuyển trên màng tế bào thực vật giữ vai trò chính trong các cơ chế chống chịu lại với biotic và abiotic stress (các stress do các nhân tố sinh học và phi sinh học gây ra), đặc biệt là các kênh Na+ và K+ liên quan đến tính trạng chống chịu mặn. Có nhiều con đường vận chuyển Na+ đi vào rễ thực vật. Na+ có thể đi xuyên màng plasma theo các kênh và thể vận chuyển chất dinh dưỡng.
Các thể đột biến liên quan đến các kênh vận chuyển làm giảm sự tích lũy Na+ trong tế bào nhưng chỉ một vài kênh khi bị đột biến mới cho thấy hiệu ứng trực tiếp trong việc hạn chế sự vận chuyển ion Na+ vào tế bào rễ. Các kênh Ca++ không chuyên biệt (calcium permeable nonselective cation channels=NSCCs) bao gồm CNGC (CYCLIC NUCLEOTIDE-GATED CHANNEL) và GLR (GLUTAMATE-LIKE RECEPTOR) cho phép Na+ đi qua và xâm nhập vào tế bào (Hình 1).
Hơn nữa, kênh vận chuyển Na+ OsHKT2;1 trên cây lúa cho thấy hỗ trợ dòng ion Na+ đi vào rễ khi rễ bị thiếu ion K+. Trên cây Arabidopsis, thể đối chuyển (antiporter) AtCHX21-cation/H+ biểu hiện trong vùng nội bì rễ có vai trò trong việc vận chuyển Na+ từ các tế bào nội bì vào vùng tủy của rễ. Na+ xâm nhập vào xylem thông qua dòng thoát ra từ các tế bào tủy và sau đó được vận chuyển đến các mô trên không chủ yếu ở lá, cành. Các ứng viên tiềm năng cho quá trình điều khiển dòng Na+ dẫn nhập vào xylem là các kênh K+ hướng ngoại KORC và NORC. Ở Arabidopsis, AtSKOR-một thể tương đồng của KORC có mối liên hệ với hiện tượng nhập K+ vào xylem.Hơn nữa, nhóm I của các thể vận chuyển HKT giữ chức năng quan trọng trong việc loại bỏ Na+ khỏi xylem.
Sự cân bằng ion của tế bào trong suốt quá trình stress mặn đòi hỏi quá trình duy trì và phân bổ nồng độ K+ ổn định giúp cho việc tích lũy K+ trong tế bào thực vật cân bằng với Na+ (sự tích lũy nhiều Na+ sẽ gây độc cho tế bào). Biểu hiện vượt mức thể vận chuyển chuyên biệt cho K+ (không chuyên chở Na+ dù cả hai ion khá tương đồng về mặt điện tích) từ cây lúa OsHAK5 trên các tế bào BY2 cây thuốc lá giúp gia tăng tính chống chịu mặn. Kênh K+ hướng nội và hướng ngoại bào đã được xác định có liên quan đến cơ chế kiểm soát dòng vận chuyển đường dài hướng vào và ra khỏi tế bào, các kênh này giúp làm giảm tính độc ion Na+.
Vai trò của NHX và SOS trong việc duy trì hàm lượng ion Na+ thấp trong tế bào chất
Hai yếu tố chính duy trì lượng Na+ thấp trong tế bào chất ở tế bào thực vật là các kênh trao đổi (exchanger) Na+/H+ hiện diện trên màng không bào (tonoplast ) NHX1 và kênh đối chuyển (antiporter) Na+/H+ có tên SOS1 trên màng sinh chất (plasma membrane). Hầu hết các kênh NHXs cần thiết cho quá trình khử độc Na+ thông qua sự cô lập Na+ trong không bào thực vật, trong khi con đường tín hiệu SOS được ghi nhận là xuất Na+ ra khỏi tế bào.
Biểu hiện vượt mức gene AtNHX1 trên Arabidopsis và các gene tương đồng trên cà chua hoặc lúa giúp làm gia tăng tính chống chịu mặn. Các nghiên cứu gần đây cho thấy rằng các protein cấu trúc giống với NHX cũng đóng vai trò quang trọng trong việc phân ngăn ion K+ vào trong không bào. Các protein này cũng tham gia vào quá trình cân bằng pH của tế bào.
Biểu hiện vượt mức AtNHX1 trên cây cà chua giúp làm gia tăng ion K+ trong không bào cũng như sự vận chuyển K+ từ rễ lên chồi, quá trình này mang lại nhiều lợi ích cho tế bào vì nó giúp gia tăng tỉ lệ K+/Na+ nội bào (nghĩa là K+ tăng, Na+ giảm) đồng thời làm giảm hiệu ứng của stress mặn do Na+ gây ra.
Hơn nữa, các bản đồ về gene LeNHX3 trên cà chua giúp xác định tính trạng QTL liên quan đến sự tích tụ Na+ ở mô lá. Các nghiên cứu gần đây đã chứng minh rằng các kênh đối chuyển (antiporter) trên màng không bào giữ nhiều vai trò trong quá trình cân bằng thẩm thấu của tế bào, sự tăng trưởng tế bào cũng như sự phát triển của toàn bộ cơ thể thực vật. Trong khi đó, các kênh đổi chuyển NHX trên endosome có vai trò thiết yếu trong quá trình tăng trưởng của tế bào và có thể có tham gia vào các hoạt động vận chuyển nội bào, chế biến protein và phân phối sản phẩm biến dưỡng của tế bào.
Các nghiên cứu cũng đánh giá vai trò của các protein vận chuyển thuộc endosome bao gồm các protein NHX tham gia vào tính chống chịu mặn ở thực vật bằng cách kiểm soát pH và sự cân bằng ion của bào quan endosome. NHX5 và NHX5 cùng hiện diện trên bộ máy Golgi và mạng lưới trans-Golgi, các cây bị bất hoạt đồng thời các gene này (knockout) nhx5 và nhx6 làm cây mẫn cảm hơn với stress mặn và giảm tính chống chịu.
Sự mất chức năng của H+-ATPase (V-ATPase) trên không bào không làm thay đổi tính chống chịu mặn trên cây Arabidopsis.Ngược lại, giảm sự hoạt động của V-ATPase trên mạng lưới trans-Golgi hoặc endosome (TGN/EE) làm gia tăng sự mẫn cảm đối với stress mặn.Sự biểu hiện vượt mức H+-PPase AVP1 loại I trên không bào giúp cải thiện tính chống chịu mặn ở thực vật thông qua quá trình cô lập Na+ trong không bào. Tiềm năng ứng dụng việc biến đổi hoạt động của H+-PPase AVP1 trên cây trồng được thể hiện qua sự biểu hiện vượt mức gene AtAVP1 trên lúa mạch (Hordeum vulgare) giúp gia tăng tính chống chịu mặn dưới điều kiện nhà lưới, bên cạnh đó cũng giúp cây tăng sinh khối và năng suất khi thử nghiểm trên các cánh đồng bị nhiễm mặn.
HKT: nhân tố chính trong sự chống chịu mặn ở thực vật và quá trình phân ngăn Na+ từ rễ lên chồi
Sự xác định gene HKT1 ở lúa mì giúp hỗ trợ quá trình vận chuyển ion Na+/K+ đã giúp khám phá ra nhiều gene HKT tương đồng ở những loài thực vật khác. Các phân tích về trình tự cũng như khả năng vận chuyển đã cho thấy ít nhất hai phân nhóm của các thể vận chuyển HKT, nhóm I và II. Trong hầu hết các trường hợp hỗ trợ quá trình vận chuyển có chọn lọc ion Na+ và sự đồng vận chuyển Na+_K+ thì cả hai nhóm này đều tham gia vào, nhóm I ứng với sự vận chuyển Na+ có chọn lọc còn nhóm II đồng vận chuyển Na+_K+. Các đột biến gián đoạn một gene HKT trên Arabidopsis (AtHKT1;1) mã hóa cho một thể vận chuyển nhóm I đã làm gia tăng sự mẫn cảm và tích tụ nhiều Na+ trên mô lá sau khi cây bị stress mặn, quá trình này cũng đi đôi với hiện tượng giảm nồng độ Na+ ở rễ.
Các phân tích xa hơn đã chứng minh một vai trò chính của gene AtHKT1;1 và gene tương đồng của nó trên lúa OsHKT1;5 trong quá trình loại bỏ Na+ khỏi dịch mộc (dòng dung dich duy chuyển trong mạch mộc từ rễ lên lá) và chuyển các Na+ vào các tế bào nhu mô mộc xung quanh, thông qua quá trình này sẽ bảo vệ mô lá khỏi độc tính của Na+. Biểu hiện vượt mức gene AtHKT1;1 trong mô tủy giúp tăng cường tính chống chịu mặn.
Các phân tích sinh lý học in vivo trên các tế bào rễ đã cho thấy gene AtHKT1;1 hỗ trợ vận chuyển Na+ thông qua các thể vận chuyển giống các kênh vận chuyển chuyên biệt theo hình thức thụ động. Sự loại bỏ Na+ khỏi mạch mộc được hỗ trợ bởi gene AtHKT1;1 cũng giúp kích thích quá trình nhập ion K+ vào mạch mộc một cách gián tiếp thông qua các kênh K+, điều này làm gia tăng tỉ lệ K+/Na+ trên lá giúp cây chống lại stress do Na+ gây ra.
Phân tích các QTL quy định tính trạng kháng lại độc tính của Na+ đã cho thấy các cơ chế tháo Na+ khỏi mạch mộc (xylem) thiết yếu cho quá trình chống chịu mặn trên lúa và lúa mì. Trong cả hai trường hợp trên, các vùng QTL chống chịu mặn đã được xác định bao gồm gene tương đồng của HKT1;5. Gene này mã hóa một thể vận chuyển HKT thuộc nhóm I có tính chọn lọc cao đối với Na+. Phân tích QTL chống chịu Na+ trên lúa mì đã dẫn đến việc phát hiện một QTL khác giúp cây chống chịu mạnh đối với stress mặn, QTL được đặt tên Nax1. QTL Nax1 đã được xác định tại vùng mã hóa của gene TaHKT1;4 mà gene này tạo ra thể vận chuyển HKT thuộc nhóm I giúp loại bỏ Na+ khỏi mạch mộc ở mô lá.
Các phân tích so sánh giữa các giống lúa indica chống chịu mặn và các giống nhiễm japonica đã đưa ra giả thuyết rằng gene OsHKT1;4 giúp giảm sự vận chuyển Na+ từ bẹ lá lên phiến lá lúa khi cây gặp stress mặn. Gần đây người ta đã sử dụng chỉ thị phân tử để đưa gene HKT1;5 từ lúa mì có họ hàng cổ đại vào lúa mì trồng thương mại giúp làm tăng năng suất lúa mì trên các đồng lúa thử nghiệm bị nhiễm mặn ở Australia. Các phát hiện trên đã giúp chứng minh rằng các thể vận chuyển HKT nhóm I hiện diện ở tế bào nhu mô của mạch mộc đóng vai trò chủ chốt trong cơ chế chống chịu mặn và bảo vệ các cơ quan đảm trách chức năng quang hợp khỏi sự tích tụ quá nhiều Na+ trong suốt quá trình stress mặn.
Sự duy trì K+ và loại bỏ Na+ ở mô lá cho thấy có mối tương quan rất mạnh đến sự chống chịu mặn ở thực vật. Điều này cũng phù hợp với các nghiên cứu về HKT nhóm I như là các yếu tố chính trong việc xác định tỉ lệ K+/Na+ ở thực vật. Tuy nhiên, nghiên cứu sâu trên các mô chồi và kỹ thuật “genome-wide association” với hơn 300 accessions đã chỉ ra các accession có sự tích tụ Na+ ở nồng độ cao ở lá sẽ có khả năng phát triển trên các vùng đất nhiễm mặn như các vùng đất ven biển. Nồng độ Na+ cao ở lá có liên quan đến một allele yếu của gene AtHKT1;1. Allele này làm giảm sự biểu hiện của AtHKT1;1 ở rễ.
Các nhà khoa học đề nghị rằng allele yếu AtHKT1;1 kiểm soát lượng Na+ có mức độ và làm giảm độc tính của Na+. Cơ chế này quan trọng trong việc điều chỉnh áp suất thẩm thấu của tế bào trong quá trình cây bị stress mặn. Allele yếu AtHKT1;1 có thể chỉ đạt lợi ích khi hiện diện trong một số cây có nền tảng di truyền mang tính chống chịu tốt. Các cây này có thể mang các cơ chế chống chịu mặn khác cần được khám phá.
Để tạo ra cây trồng chống chịu mặn thì nhiều kiến thức cần được nắm vững về các hoạt động tương hỗ và kết hợp của các tính trạng chống chịu (tương tác của các gene chống chịu). Một yêu cầu hiểu biết giống như vậy về việc gia tăng tích tụ Na+ ở chồi có mối tương quan với tính chống chịu mặn ở thực vật cũng đã được ghi nhận trong một nghiên cứu về HKT nhóm II ở lúa mạch. Gia tăng hấp thu Na+ ở rễ và nồng độ Na+ cao ở dịch mạch mộc (xylem sap) làm giảm sự biểu hiện của HvHKT2;1. Khác biệt với lúa và lúa mì, ở lúa mạch tỉ lệ chuyển vị Na+ cao hơn đến chồi và bao gồm cả nồng độ muối cao có thể mang một ý nghĩa quan trọng trong chiến lược kháng lại stress mặn.
Các cơ chế điều hòa biểu hiện của gene AtHKT1;1 gần đây đã được xác định. Hormone cytokinin điều hòa âm tính đối với sự biểu hiện của gene AtHKT1;1 ở rễ cây Arabidopsis theo các yếu tố điều hòa đáp ứng dạng B là ARR1 và ARR12. Do đó, trong các đáp ứng với stress mặn thì khi lượng cytokinin giảm sẽ làm tăng sự biểu hiện của gene AtHKT1;1. Có một cơ chế điều hòa âm tính gene AtHKT1;1 ở rễ Arabidopsis cũng đã được ghi nhận trong một nghiên cứu gần đây, đó là sự điều hòa âm tính của yếu tố phiên mã ABI4 (ABA-INSENSITIVE 4). Đột biến mất chức năng của gene ABI4 làm cây gia tăng tính chống chịu mặn với sự biểu hiện lượng Na+ thấp ở chồi tương ứng với gia tăng sự biểu hiện gene AtHKT1;1. Ngược lại, khi biểu hiện vượt mức gene ABI4 ở các cây biểu hiện kém gene AtHKT1;1 làm cây mẫn cảm hơn với stress mặn.
Nhiều yếu tố điều hòa cis tại khu vực promoter của gene AtHKT1;1 đã được xác định khi sàn lọc các yếu tố kìm hãm ở cây đột biến gene sos3. Một đoạn DNA chèn vào vùng trình tự lặp lại theo kiểu “tandem” với kích thước khoảng hơn 3.9kb vùng upstream mã khởi đầu (start codon) của gene AtHKT1;1 đã dẫn đến sự giảm biểu hiện gene AtHKT1;1 rõ rệt trên rễ cây đột biến gene sos3. Trình tự lặp lại kiểu “tandem” đã được đề nghị như là yếu tố tăng cường (enhancer element) trong quá trình điều hòa sự biểu hiện của gene AtHKT1;1 trong khi đó thì người ta vẫn chưa xác định được yếu tố phiên mã nào tham gia vào việc tăng cường sự biểu hiện của gene này.
Sàn lọc di truyền đột biến gene sss (soil salinity sensitive) đã giúp xác định được gene sss1-1 đột biến cho thấy kiểu hình rất nhạy cảm với ion Na+ trên chồi.Gene SSS1 mã hóa cho protein mang tên AtrbohF, đây là một enzyme NADPH oxidase tham gia xúc tác tạo các phần tử ROS (reactive oxygen species). Sự thiếu vắng protein AtrbohF ở vùng chu luân của rễ (pericycle) và nhu mô mạch đã làm giảm sự tích tụ ROS do stress mặn gây ra và cũng làm gia tăng quá mức Na+ trên chồi cũng như trong dịch mộc.
Sự sản xuất ROS được hỗ trợ bởi protein AtrbohF có thể tăng cường tháo Na+ khỏi dịch mộc mà quá trình này do gene AtHKT1;1 tham gia kiểm soát. Hiện tượng này giúp bảo bệ mô lá khỏi stress mặn, tuy nhiên nhiều nghiên cứu khác cần được thực hiện để kiểm tra mô hình giả thuyết này hoặc các cơ chế khác có thể tham gia vào quá trình kiểm soát tính chống chịu mặn. Với các kết quả đạt được vừa kể trên có thể cung cấp các chứng cứ để ủng hôn giả thuyết rằng sự biểu hiện của gene AtHKT1;1 được kiểm soát bởi một mạng lưới thông tin tín hiệu phức tạp qua nhiều bước với nhiều yếu tố điều hòa cis-trans khi cây bị stress mặn.
Sự biến đổi của chromatin và di truyền biểu sinh trong sự chống chịu mặn ở thực vật
Sự biến đổi chromatin được biết như sự biến đổi mang tính di truyền biểu sinh giúp thực vật thích nghi với các điều kiện môi trường stress khác nhau. Nhiều nghiên cứu đã chỉ ra rằng sự biến đổi chromatin xuất hiện trong quá trình thực vật đáp ứng với stress mặn trong cùng một thế hệ. Xét theo khía cạnh này, khi cây được xử lý trong môi trường stress mặn ở nồng độ thấp thì khi chuyển sang môi trường bình thường để cây tăng trưởng, sau đó khi gặp stress mặn nồng độ cao thì cây sẽ thích nghi tốt hơn (chống chịu tốt hơn).
Với thí nghiệm trên, sau khi cây chuyển từ môi trường stress mặn với nồng độ muối thấp và chuyển sang môi trường phát triển bình thường thì người ta không thấy sự khác biệt về kiểu hình của cây xử lý stress mặn thấp so với cay không xử lý. Sau đó các cây được chuyển tiếp sang môi trường có nồng độ muối cao hơn thì các cây đã được xử lý stress mặn nhẹ sẽ tích tụ ít ion Na+ hơn so với cây không xử lý và do đo sẽ chống chịu mặn tốt hơn.
Hiện tượng trên có thể quy cho quá trình biến đổi histone trên chromatin mang tính di truyền biểu sinh mà quá trình này ảnh hưởng đến sự biểu hiện của các yếu tố phiên mã (transcription factor). Người ta cũng thấy rằng gene AtHKT1;1 biểu hiện mạnh hơn ở các cây được xử lý stress mặn nhẹ so với cây không xử lý, điều này có thể giả thích cho việc thay đổi quá trình tích tụ Na+. Các nghiên cứu khác cho thấy sự methyl hóa amino acid cytosine không được thực hiện trên vị trí đích giả định của RNA kích thước nhỏ thuộc vùng promoter của gene AtHKT1;1 đã dẫn tới việc làm giảm sự biểu hiện của gene này và gây ra sự mẫn cảm đối với stress mặn.
Các nghiên cứu xa hơn trên cây đậu nành cho thấy stress mặn ảnh hưởng đến sự methyl hóa DNA tại nhiều vùng promoter và các vùng mã hóa của 4 yếu tố phiên mã. Điều này chỉ ra rằng sự biến đổi của chromatin nằm trong vùng hiện diện của các gene này có thể tăng cường tính chống chịu mặn cho cây.
Sự methyl hóa cũng như sự biểu hiện của các gene có chức năng làm biến đổi chromatin khác nhau giữa các giống lúa cũng như các mô hay cơ quan của cây dưới điều kiện stress mặn. Cho đến hiện tại, các nghiên cứu về stress mặn đã không cho thấy một cách rõ ràng về di truyền biểu sinh của sự chống chịu mặn từ thế hệ này sang thế hệ khác. Một yếu tố khác cũng quan trọng khi nghiên cứu về stress mặn cũng cần được lưu ý là trạng thái đa bội thể ở thực vật. Sự tự đa bội hóa ở thực vật gần đây cho thấy rằng có thể tham gia vào sự chống chịu mặn ở nồng độ muối cao và hiện tượng này cũng có mối liên quan đến quá trình tích lũy ion K+ hiệu quả trên cây Arabidopsis.
Vai trò và tầm quan trọng của các phân tử điều hòa thẩm thấu (osmolyte)
Sự tích tụ các osmolyte hữu cơ như proline, glycine betaine, sugar alcohol, polyamine và các protein họ LEA (late embryogenesis abundant) giữ vai trò quan trọng trong quá trình duy trì thế thẩm thấu thấp trong tế bào thực vật cũng như tham gia vào việc ngăn chặn các ảnh hưởng bất lợi của stress mặn. Các nghiên cứu ở mức phân tử cho thấy stress mặn kích hoạt sự tổng hợp proline và sự phân hủy phân tử này được tăng cường trong suốt giai đoạn hồi phục của cây. Trong suốt giai đoạn hồi phục này của cây, proline đóng vai trò như một phân tử tín hiệu và được xem có vai trò điều hòa sự phân bào, sự chết tế bào cũng như điều hòa biểu hiện các gene tham gia vào giai đoạn hồi phục sau stress.
Trên cây Arabidopsis, khi bất hoạt gene P5CS1 mã hóa cho enzyme tổng hợp ∆-1-pyrroline-5-carboxylate đã làm giảm sự tổng hợp proline làm cây siêu mẫn cảm đối với stress mặn. Trong nhiều năm người ta cho rằng proline đóng vai trò quan trọng trong quá trình điều hòa thẩm thấu của tế bào, tuy nhiên nhiều ý kiến khác lại cho rằng nó hoạt động như là yếu tố chống lại các phân tử mang gốc oxy hóa mạnh (như các ROS, AOS), chất đệm trong phản ứng oxy hóa khử của tế bào, hoặt có vai trò như là một chaperone, tham gia ổn định cấu trúc protein và cấu trúc màng tế bào dưới các điều kiện stress.
Tương tự như proline, glycine betain là một osmolyte hữu cơ được tổng hợp trên rất nhiều loài thực vật để tạo ra sự cân bằng thế thẩm thấu của tế bào dưới điều hiện stress mặn. Có chứng cứ cho thấy rằng glycine betaine là một chất tan tham gia vào quá trình bảo vệ các enzyme chủ chốt và cấu trúc màng tế bào. Mặc dù glycine betaine được ghi nhận đóng vai trò quan trọng trong việc duy trì hoạt động của của enzyme dọn dẹp các ROS nhưng vẫn chưa thấy chứng cứ chứng minh liệu rằng glycine betaine hoặc các phân tử khác tham gia trực tiếp vào các hoạt động khử các yếu tố ROS.
Tạo ra cây trồng chống chịu mặn
Một ví dụ về biến đổi di truyền cho cây chống chịu mặn được thực hiện trên yếu tố phiên mã được cảm ứng bởi stress mặn bZIP24. Yếu tố phiên mã này cảm ứng sự biểu hiện của rất nhiều gene liên quan đến đáp ứng stress mặn ở cây Arabidopsis. Bất hoạt gene bZIP24 bằng kỹ thuật RNAi (RNA interference) cho thấy gia tăng sự chống chịu mặn của cây. Điều này có thể giải thích là do gene AtHKT1;1 là một trong những gene mục tiêu bị bZIP24 ức chế sự biểu hiện thông qua cách trực tiếp hoặc gián tiếp.
Ban đầu bZIP24 được khám phá trong một nghiên cứu so sánh về sự điều hòa phiên mã trên cây Arabidopsis và loài chống chịu mặn cải gió ngọt (sweet alyssum).Đây là ví dụ điển hình về sử dụng loài cây chịu mặn và nghiên cứu so sánh genomics trong việc khám phá ra các có chế chống chịu mặn.
Sự biểu hiện của gene AtHKT1;1 giữ vai trò quan trọng trong việc xác định các mức Na+ trên lá thể hiện tính chống chịu mặn. Cơ chế phân tử điều hòa biểu hiện gene AtHKT1;1 và hoạt động của nó có thể là yếu tố quan trọng phục vụ cho việc nghiên cứu tạo ra cây trồng kháng mặn. Tuy nhiên bên cạnh các yếu tố điều hòa âm tính đã được đề cập bên trên thì các yếu tố điều hòa dương tính vẫn chưa được khám phá nhiều thông tin hữu ích.
Ngoài các nghiên cứu trên cây mô hình Arabidopsis, các nghiên cứu trên các cây trồng khác cũng đã phát hiện thêm những thông tin giá trị về cơ chế chống chịu mặn ở thực vật. Yếu tố phiên mã mang tên SERF1 (SALT-RESPONSIVE ERF1) đã được xác định như là một yếu tố tăng cường hoạt động của chuỗi phản ứng enzyme MAP KINASE được kích hoạt bởi ROS trong suốt quá trình stress mặn. Sự biểu hiện của SERF1 được cảm ứng trên rễ do nồng độ muối cao, các cây lúa bị mất chức năng của yếu tố SERF1 cho thấy khống có sự biểu hiện của các gene tham gia giúp cây chống chịu lại stress mặn.
Các cây lúa đột biến gene serf1 đạt 3-4 tuần tuổi được trồng trên hệ thống thủy canh cho thấy không chịu được stress mặn. Trong khi đó, các cây lúa biểu hiện vượt mức gene SERF1 cho thấy sự gia tăng tính chống chịu mặn. Các phân tích cho thấy ở cây lúa đột biến serf1 tỉ lệ Na+/K+ ở lá gia tăng một cách có ý nghĩa so với cây bình thường.
MAS như là công cụ lý tưởng trong nghiên cứu chọn giống
Cải tiến năng suất cây trồng là mục tiêu không ngừng nghĩ của các chương trình nghiên cứu chọn tạo giống. Chọn giống chống chịu mặn theo phương pháp truyền thống thường mất thời gian rất lâu và các phương pháp mới luôn được tìm tòi để hỗ trợ hoặc thay thế cho các phương pháp cũ. Thông thường, các giống cây trồng được lai tạo từ các nguồn vật liệu mang tính đa dạng di truyền, sau đó được chọn lọc dựa trên kiểu hình biểu hiện ra bên ngoài trong suốt quá trình tăng trưởng và phát triển. Quá trình chọn lọc này có thể được thực hiện trong phòng thí nghiệm hay ngoài đồng.
Quá trình cải tiến và hỗ trợ phương pháp chọn giống truyền thống hiện tại sử dụng các phân tích QTL và chọn lọc dựa vào marker (MAS).MAS là một phương pháp quan trọng và hiệu quả yêu cầu sự liên kết giữa một tính trạn mục tiêu và marker di truyền mà các marker này mang tính đa dạng di truyền giữa các dòng bố mẹ sử dụng làm vật liệu lai. Do đó, để áp dụng MAS thành công trong công tác nghiên cứu chọn giống thì phải có kiến thức chuyên sâu về di truyền các tính trạng mục tiêu và sự biến dị trên các loài cây trồng mục tiêu.
Một ví dụ tiêu biểu trong chọn giống cây trồng chống chịu mặn là trường hợp QTL Saltol, một QTL được xác định trên lúa giúp cây chống chịu điều kiện stress mặn. Các vùng genome khác cũng cho thấy vai trò quan trọng trong cơ chế chống chịu mặn bên cạnh QTL Saltol.Các marker phân tử và các kỹ thuật giải trình tự thế hệ mới sẽ đóng góp vai trò quan trọng trong chiến lược chọn giống trong tương lai.
MAS rõ ràng là một công cụ mạnh mẽ và hiệu quả trong việc nghiên cứu chọn tạo giống cây trồng kháng mặn. Một điểm quan trọng là việc chọn lựa các marker thích hợp cho chương trình chọn giống chống chịu mặn.Có nhiều nghiên cứu đã cố gắng cải thiện khả năng chống chịu mặn của thực vật thông qua việc biến đổi di truyền bằng cách thay đổi một số gene nhất định.Tuy nhiên, các gene này không cho thấy hiệu quả để cải thiện khả năng sống sót một cách có ý nghĩa của cây trồng khi gặp môi trường có nồng độ muối cao.
Người ta đề xuất rằng để gia tăng tính chống chịu mặn trên cây trồng chỉ cẩn thay đổi một hoặc vài thành phần chính hệ thống gene điều hòa trong cơ chế chống chịu mặn thay vì thay đổi rất nhiều cơ chế phân tử liên quan đến tính chống chịu mặn. Bên cạnh phương pháp chọn giống truyền thống thì phương pháp chồng gene dựa vào marker phân tử cũng hứa hẹn là một phương pháp hữu ích trong việc cải thiện tính chống chịu mặn ở cây trồng.Tuy nhiên, phương pháp chồng gene gặp trở ngại là do sự phân li độc lập của các tính trạng mục tiêu với nhau, điều này làm cho công việc chọng giống thêm khó khan và phức tạp.
Một phương pháp mới nổi lên gần đây trong chiến lược chọn tạo giống cây trồng kháng mặn để tránh vấn đề vừa nêu là sử dụng kỹ thuật biến đổi genome trực tiếp thông qua các kỹ thuật sử dụng các enzyme nuclease như các kỹ thuật ZFN, TALEN và CRISPR-Cas9.